

Southern blot
Nguyên tắc của Southern blot là màng lai nitrocellulose có khả năng tiếp nhận DNA đã được biết từ lâu và đã được sử dụng trong các nghiên cứu lai axit nucleic khác nhau vào những thập niên 1950 và 1960. Vào thời kỳ này DNA cố định không được phân đoạn, chỉ đơn giản bao gồm DNA tổng số được gắn trên màng lai nitrocellulose. Sự ra đời của phương pháp điện di trên gel vào đầu thập niên 1970 đã cho phép các đoạn DNA được cắt bởi enzyme hạn chế có thể được phân tách dựa trên cơ sở kích thước của chúng. Từ đó bước phát triển tiếp theo của phương pháp là chuyển các đoạn DNA phân tách từ gel lên màng lai nitrocellulose. Phương pháp này được E. M. Southern mô tả tại Ðại học Edingburgh vào năm 1975. Phương pháp Southern blot đơn giản và hiệu quả. Mặc dù đã được cải tiến nhưng phương pháp đang được sử dụng ở nhiều phòng thí nghiệm sinh học phân tử sai khác không đáng kể so với phương pháp ban đầu.
Southern blot bao gồm các bước cơ bản sau:
-Cắt DNA bằng enzyme hạn chế thích hợp.
-Ðiện di sản phẩm cắt trên gel agarose.
-Làm biến tính DNA (thông thường khi nó còn ở trên gel): ví dụ có thể nhúng nó vào trong dung dịch NaOH 0.5M, DNA sợi kép sẽ được tách thành DNA sợi đơn. Chỉ DNA sợi đơn mới có thể chuyển lên màng lai.
-Chuyển DNA đã biến tính lên màng lai. Thông thường màng lai được sử dụng là màng nitrocellulose. Người ta cũng có thể sử dụng màng nylon. Màng nitrocellulose điển hình có khả năng tiếp nhận 100µg DNA/cm2, trong khi màng nylon có khả năng tiếp nhận 500µg DNA/cm2. Mặt khác màng nylon có khả năng giữ DNA chắc hơn và ít đứt gãy hơn. Việc chuyển DNA thường được tiến hành bằng hoạt tính mao dẫn trong khoảng vài tiếng hoặc có thể dùng một thiết bị thấm chân không. Nếu dùng thiết bị thấm chân không thì sẽ nhanh hơn, chỉ mất khoảng một tiếng. Trong quá trình chuyển, vị trí các đoạn DNA vẫn được giữ nguyên không thay đổi.
-Lai DNA đã được cố định trên màng với mẫu dò (probe) DNA có đánh dấu. Quá trình này dựa trên nguyên tắc bổ sung (giữa DNA trên màng lai với mẫu dò). Ðể đánh dấu người ta thường sử dụng P32, biotin/streptavidin hoặc một mẫu dò phát quang sinh học.
-Ðịnh vị các phân tử lai DNA-mẫu dò. Nếu sử dụng mẫu dò đánh dấu phóng xạ thì dùng phương pháp phóng xạ tự ghi (autoradiograph) để xác định, nếu sử dụng biotin/streptavidin thì dùng phương pháp so màu hoặc nếu sử dụng mẫu dò phát quang sinh học thì phát hiện bằng sự phát quang.
Phương pháp Southern blot được thiết kế để xác định sự hiện diện, kích thước, số lượng bản sao, tính đồng dạng của DNA trong một phức hợp. Ví dụ, Southern blot có thể được sử dụng để phát hiện một gen đặc biệt ở trong một genome nguyên vẹn.
Northern blot
Sau khi E. M. Southern mô tả phương pháp Southern blot vào năm 1975, người ta đã dùng một phương pháp tương tự để xác định các đoạn RNA đặc biệt gọi là Northern blot. Phương pháp này còn được gọi là Northern blotting, phương pháp lai Northern hay phương pháp lai RNA.
Northern blot bao gồm các bước cơ bản sau:
-RNA (RNA tổng số hoặc chỉ mRNA) được phân tách bằng điện di trên gel agarose.

Sơ đồ mô tả phương pháp Southern blot
-RNA sau khi đã phân tách được chuyển lên màng lai (các phân tử RNA giữ nguyên vị trí như ở trên gel).
-RNA cố định trên màng được lai với mẫu dò DNA sợi đơn (hoặc RNA) có đánh dấu phóng xạ hoặc được gắn với một enzyme (alkalin phosphatase hoặc horseradish peroxidase) tạo thành phân tử lai RNA-DNA (hoặc RNA-RNA) sợi kép.

Sơ đồ mô tả phương pháp Northern blot
-Vị trí của mẫu dò được phát hiện nhờ kỹ thuật phóng xạ tự ghi nếu nó được đánh dấu phóng xạ. Trong trường hợp mẫu dò được gắn với enzyme thì đem ủ với một cơ chất không màu. Enzyme liên kết với nó sẽ biến đổi thành một sản phẩm màu có thể nhìn thấy hoặc phát ra ánh sáng mà sẽ được phát hiện bằng phim X quang một cách trực tiếp.
Phương pháp Northern blot cho phép phát hiện sự có mặt, xác định kích thước, trọng lượng phân tử, khối lượng mRNA ở trong các mẫu khác nhau. Ðây là một phương pháp rất tốt để phân tích sự biểu hiện của gen khi chúng ta cần định lượng để phân biệt sự khác nhau giữa hai mẫu và nó rất nhạy bởi vậy chúng ta có thể điện di một lượng lớn RNA tổng số hoặc mRNA trên gel.
Western blot
Western blot là phương pháp có độ nhạy cao dựa trên tính đặc hiệu của kháng thể để phát hiện protein đã được điện di trên gel SDS-PAGE (sodium dodecyl sulfate-polyacrylamide gel electrophoresis) và chuyển lên màng lai.
Western blot cho phép xác định sự có mặt, trọng lượng phân tử, định lượng protein có mặt trong các mẫu khác nhau.
Western blot còn có tên gọi khác là Western blotting hay là phương pháp lai thấm protein.
Western blot bao gồm các bước cơ bản sau:
-Protein được phân tách bằng điện di trên gel SDS-PAGE.
-Các protein được chuyển sang màng lai nitrocellulose, giữ nguyên vị trí như đã phân tách trên gel.
-Ủ màng lai đã cố định protein với một kháng thể sơ cấp (primary antibody). Kháng thể sơ cấp là một kháng thể đặc hiệu, sẽ bám vào protein và tạo thành một phức hợp protein-kháng thể đối với protein quan tâm.
-Tiếp theo ủ màng lai với một kháng thể thứ cấp (secondary antibody) có enzyme (alkalin phosphatase hoặc horseradish peroxidase) đi kèm. Kháng thể thứ cấp sẽ bám vào kháng thể sơ cấp giống như kháng thể sơ cấp đã bám vào protein.
-Tiếp tục ủ màng lai trong một hỗn hợp phản ứng đặc hiệu với enzyme. Nếu mọi việc đều diễn ra một cách chính xác sẽ phát hiện thấy các băng ở bất kỳ nơi nào có mặt phức hợp protein-kháng thể sơ cấp- kháng thể thứ cấp-enzyme hay nói cách khác là ở bất kỳ nơi nào có mặt protein quan tâm.
-Ðặt một phim nhạy cảm với tia X lên màng lai để phát hiện các điểm sáng phát ra do enzyme.

Sơ đồ mô tả phương pháp Western blot
ELISA (Enzymee-Linked Immunosorbent Assay)
ELISA được mô tả lần đầu tiên vào năm 1971 và từ đó đã trở thành một phương pháp được sử dụng ngày càng rộng rãi và quan trọng hơn trong nghiên cứu, chẩn đoán và xét nghiệm bởi vì nó có khả năng phát hiện nhạy bén với một lượng vật chất rất nhỏ.
ELISA đã thay thế một số kỹ thuật huyết thanh “cổ điển“ phức tạp, cồng kềnh tốn nhiều thời gian hơn và mở rộng phạm vi phương pháp phát hiện virus cũng như các marker liên quan đến sự nhiễm của chúng.
Xét nghiệm ELISA có thể được tiến hành với một số phương pháp như ELISA “trực tiếp“, “gián tiếp“, “sandwich“ và “cạnh tranh“.
Nguyên tắc cơ bản của phương pháp ELISA là kháng nguyên đã hoà tan trong dung dịch đệm thích hợp có thể phủ lên bề mặt plastic (như polystyrene). Quá trình này có thể là trực tiếp hoặc thông qua một kháng thể. Khi huyết thanh được thêm vào, các kháng thể có thể kết hợp với kháng nguyên ở pha đặc (solid phase).
Xét nghiệm ELISA được thực hiện trong đĩa plastic kích thước 8cm x 12cm, chứa 8x12 giếng. Mỗi giếng có chiều cao khoảng 1cm và đường kính là 0,7cm (Hình dưới).

Ðĩa plastic sử dụng để tiến hành xét nghiệm ELISA
Các kháng thể sử dụng trong phương pháp ELISA được gắn với enzyme bằng liên kết đồng hoá trị. Kháng nguyên được gắn với giếng plastic và kháng thể liên kết với enzyme được gắn với kháng nguyên. Kháng thể không gắn kháng nguyên sẽ bị rửa trôi đi. Enzyme được giữ lại và vì vậy lượng kháng thể gắn enzyme được phát hiện bằng cách cho thêm vào một cơ chất làm thay đổi màu do hoạt tính của enzyme. Ðộ màu tạo thành là tỉ lệ với lượng enzyme bám ở giếng plastic, từ đó suy ra lượng kháng thể, sau đó tiếp tục suy ra lượng kháng nguyên (Hình dưới).
Tính nhạy của ELISA là do sự khuyếch đại bởi hoạt tính enzyme. Mỗi một phân tử enzyme bám vào kháng thể có thể tạo ra hàng ngàn phân tử màu do hoạt tính enzyme. Trước khi các kháng thể gắn enzyme có thể được sử dụng rộng rãi, các kháng thể phóng xạ đã được sử dụng trong kỹ thuật miễn dịch phóng xạ (radio immuno assays-RIA). Kỹ thuật RIA như là một đột phá có ý nghĩa và người sáng tạo ra nó là Rosalyn Yalow đã được nhận giải thưởng Nobel Sinh lý và Y học vào năm 1977.

Nguyên tắc của phương pháp ELISA
Phương pháp PCR
Phương pháp PCR (polymerase chain reaction-phản ứng tổng hợp dây chuyền nhờ polymease) là một trong những phương pháp được sử dụng rộng rãi nhất trong lĩnh vực Sinh học phân tử. Phương pháp này do Kary Mullis phát minh vào năm 1985 và được giới thiệu lần đầu tiên tại Hội thảo lần thứ 51 ở Cold Spring Harbor vào năm 1986 và ông đã nhận được giải thưởng Nobel Hoá sinh học vào năm 1993.
Phương pháp PCR cho phép tổng hợp rất nhanh và chính xác từng đoạn DNA riêng biệt. Ðây thực sự là phương pháp hiện đại và thuận tiện cho việc xác định sự có mặt của một gen nào đó trong tế bào với độ chính xác cao.
Phương pháp này dựa trên sự khám phá hoạt tính sinh học ở nhiệt độ cao của DNA polymerase được tìm thấy trong các sinh vật ưa nhiệt (vi khuẩn sống trong các suối nước nóng). Phần lớn các DNA polymerase chỉ làm việc ở nhiệt độ thấp. Nhưng ở nhiệt độ thấp, DNA xoắn chặt vì vậy DNA polymerase không có nhiều khả năng làm biến tính phần lớn các phần của phân tử. Nhưng các polymerase chịu nhiệt này hoạt động ở nhiệt độ rất cao, có thể lên đến 100oC. Ở nhiệt độ này DNA (dạng thẳng) sẽ bị biến tính.
Các thành phần chủ yếu của phản ứng PCR
DNA mẫu (DNA template)
Ðây là mẫu DNA sinh học mà chúng ta muốn khuyếch đại. Phản ứng PCR tối ưu xảy ra trên DNA thật tinh sạch nhưng phản ứng PCR vẫn cho kết quả tốt với DNA thu nhận trực tiếp từ dịch chiết tế bào. Lượng mẫu DNA sử dụng có khuynh hướng giảm khi sử dụng các enzyme DNA polymerase cho hiệu quả cao (<100ng). Lượng DNA mẫu nếu cao quá phản ứng PCR sẽ không xảy ra. PCR còn cho phép khuyếch đại cả những mẫu DNA không được bảo quản tốt, các mẫu DNA đã bị phân hủy từng phần như ở các vết máu để lâu ngày, tinh dịch đã khô, tóc, móng tay của người chết....
Mồi (primer)
Mồi là những đoạn DNA sợi đơn ngắn và cần thiết cho việc xúc tiến phản ứng dây chuyền tổng hợp DNA . Chúng nhận ra phần DNA cần được nhân lên, bắt cặp bổ sung với một đầu của DNA mẫu và tạo ra vị trí bắt đầu tái bản. Các mồi này có chiều ngược nhau, bao gồm một mồi xuôi (forward primer) và một mồi ngược (reverse primer).
Mồi là yếu tố quan trọng nhất của phản ứng PCR, quyết định hiệu quả của phản ứng. Do đó việc thiết kế mồi cần được tuân thủ một số nguyên tắc nhất định:
+ Cả hai mồi trong một phản ứng PCR phải có nhiệt độ nóng chảy (Tm) gần như nhau bởi vì chúng được ủ ở cùng một nhiệt độ.
+ Kích thước tối thiểu của mồi là 18 base (thông thường là 18-24 base) để quá trình lai xảy ra tốt hơn.
+ Mồi phải đặc hiệu có nghĩa là nó phải đặc trưng cho trình tự DNA cần được khuyếch đại, một mồi chỉ bám vào một vị trí nhất định trên gen.
+ Không có nút cài tóc (hairpin loop): trong mỗi một mồi cần tránh trình tự đối xứng bậc hai (palindromic sequences) có thể làm tăng cấu trúc sợi bên trong ổn định do đó hạn chế việc mồi bám vào DNA mẫu .

Sự hình thành nút cài tóc do mồi chứa trình tự đối xứng bậc hai
+ Tránh sự bổ sung giữa hai mồi: sự bổ sung giữa hai mồi sẽ làm tăng sản phẩm primer dimer .

Sự bổ sung giữa hai mồi tạo nên primer dimer
+ Trật tự các base cũng ảnh hưởng đến sự ổn định việc bám của mồi dưới nhiệt độ cao. Hai trong ba base ở đầu 3’ của mồi nên là G hoặc C, vì G và C có 3 liên kết hydro do đó sự polymer hoá sẽ tốt hơn.
+ Khoảng cách tối ưu giữa hai mồi: đây là một ứng dụng rất đặc trưng, nhưng đối với phần lớn các thử nghiệm PCR chẩn đoán, tốt nhất khoảng cách giữa hai mồi khi đã bám vào DNA khuôn là 150-500 base.
Enzyme polymerase chịu nhiệt
Vào thập niên 1960, nhà Vi sinh vật học Thomas Brock đã đến Công viên Quốc gia Yellowstone (Bang Wyoming, Mỹ) để nghiên cứu các vi sinh vật ưa nhiệt sống trong suối nước nóng 80-95 oC. Ông đã phát hiện một loài vi khuẩn phát triển mạnh ở nhiệt độ cao, có tên là Thermus aquaticus. Hai mươi năm sau, các nhà khoa học của tập đoàn Cetus (Tập đoàn Công nghệ Sinh học California) đã nhận thấy rằng DNA polymerase từ Thermus aquaticus (Tag-polymerase) có khả năng giải quyết vấn đề của enzyme biến tính sau mỗi chu kỳ. DNA polymerase chịu nhiệt sử dụng cho phản ứng PCR lần đầu tiên được bán trên thị trường là Tag-polymerase.
Từ đó đến nay, một số vi sinh vật chịu nhiệt khác đã được khám phá và người ta đã tách chiết thêm được các DNA polymerase chịu nhiệt để sử dụng cho phản ứng PCR như Vent- polymerase (Tli-polymerase), Pfu- polymerase, rTth...Các hoạt tính của chúng được trình bày ở bảng dưới đây.
Các loại nucleotid
Trong phản ứng PCR, bốn loại nucleotid thường được sử dụng ở dạng deoxynucleotid: dATP, dCTP, dGTP, dTTP với nồng độ cân bằng trong một phản ứng (200μM/loại nucleotid).
Nước
Nước sử dụng cho phản ứng PCR phải thật tinh khiết, không chứa ion nào, không chứa DNAase, RNAase, enzyme cắt hạn chế... Nói cách khác là không chứa bất kỳ một thành phần nào khác.
Dung dịch đệm
Dung dịch đệm 10X (100mM KCl, 100mM (NH4)2SO4, 200mM Tris-Cl pH 8.8, 20mM MgSO4, 1% (w/v) Triton X-100)
Ion Mg2+
Nồng độ ion Mg2+ cũng là một yếu tố ảnh hưởng mạnh đến phản ứng PCR và nó tuỳ thuộc vào từng phản ứng. Nồng độ ion Mg2+ tối ưu là 150-200 μM. Người ta thấy rằng nếu nồng độ DNA quá cao thì enzyme polymerase sẽ gây ra nhiều lỗi hơn.
Ba giai đoạn trong một chu kỳ của phản ứng PCR
Có 3 giai đoạn chính trong phản ứng PCR và chúng được lặp đi lặp lại nhiều lần (chu kỳ) (thường từ 25 đến 75 chu kỳ).
Giai đoạn biến tính (denaturation)
Trong giai đoạn này phân tử DNA mẫu bị biến tính ở nhiệt độ cao (thường là từ 94-95 oC, lớn hơn nhiệt độ nóng chảy của phân tử) trong vòng 30 giây đến 1 phút, tất cả các liên kết hydro giưã hai mạch của phân tử bị bẻ gãy và tạo thành các DNA sợi đơn.
Giai đoạn lai (hybridization)
Nhiệt độ được hạ thấp ( thường từ 40-70 oC, thấp hơn nhiệt độ nóng chảy của mồi được sử dụng khoảng từ 3-5 oC) cho phép các mồi bám vào các phân tử DNA sợi đơn, đánh dấu phần DNA cần được khuyếch đại. Giai đoạn này kéo dài từ 30 giây đến một phút (còn được gọi là giai đoạn ủ).
Nếu nhiệt độ quá thấp thì các mồi sẽ gây nên nhiều lỗi và kết quả là sẽ tạo nên nhiều sản phẩm phụ. Nếu nhiệt độ quá cao thì phản ứng sẽ không có kết quả.
Công thức để xác định nhiệt độ nóng chảy (Tm) một cách tương đối là Tm=4(G+C) + 2(A+T)
Giai đoạn kéo dài (elongation)

Ba giai đoạn trong một chu kỳ của phản ứng PCR
Nhiệt độ được tăng lên đến 72oC giúp cho DNA polymerase xúc tác tổng hợp DNA tốt nhất. Công việc của DNA polymerase là di chuyển dọc theo DNA sợi đơn và sử dụng nó làm khuôn để tổng hợp sợi DNA mới bổ sung với DNA mẫu bằng cách kéo dài các phần đã được đánh dấu bởi các mồi. Thời gian của giai đoạn này phụ thuộc vào kích thước của DNA mẫu, thường kéo dài từ 30 giây đến nhiều phút.

Ðồ thị biễu diễn mối quan hệ giữa thời gian và nhiệt độ trong một chu kày của phản ứng PCR
Ở giai đoạn này của chu kỳ cuối cùng, thời gian được tăng thêm vài phút để các sợi DNA chưa được sao chép xong hoàn thành qúa trình tổng hợp.
Sau mỗi chu kỳ, số bản sao của DNA mẫu lại được tăng gấp đôi. Ðây là sự nhân bản theo cấp số nhân. Như vậy cứ 1 phân tử DNA mẫu, sau phản ứng PCR với n chu kỳ sẽ tạo thành 2n bản sao phân tử DNA
Ưu điểm của phương pháp PCR là chỉ cần một thời gian ngắn đã cho một số lượng DNA theo mong muốn.
Sản phẩm của phản ứng PCR được phân tách trên gel agarose đã nhuộm ethimidium bromide và quan sát dưới máy chiếu tia UV.
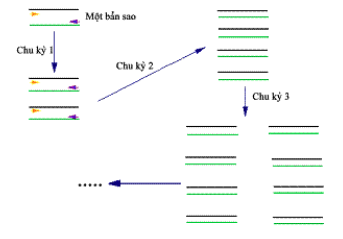
Phản ứng PCR với lượng sản phẩm tăng theo cấp số nhân
Phương pháp RT-PCR
Taq-polymerase không có hoạt tính với RNA vì vậy phản ứng PCR không thể sử dụng để khuyếch đại RNA một cách trực tiếp. Tuy nhiên có thể sử dụng kết hợp enzyme sao chép ngược (reverse transcriptase-RT) với PCR để khuyếch đại RNA. Phản ứng này sử dụng khả năng của enzyme sao chép ngược để sao chép RNA thành DNA bổ sung sợi đơn và từ DNA bổ sung sợi đơn thành DNA bổ sung sợi kép. Sau đó DNA bổ sung sợi kép sẽ được khuyếch đại nhờ Taq-polymerase.
Sản phẩm của phản ứng RT-PCR là các phân tử DNA sợi kép tương ứng với phân tử RNA khuôn mẫu. RNA tế bào tổng số tách chiết từ mô hoặc tế bào được sử dụng làm khuôn mẫu cho phản ứng sao chép ngược.
Ưu điểm của phương pháp này là cho phép nghiên cứu các mRNA với hàm lượng rất thấp mà các phương pháp như Northern blot... không thể thực hiện được.
PCR được sử dụng rộng rãi trong nhiều lĩnh vực như tạo dòng phân tử, kỹ thuật di truyền, phát hiện chẩn đoán các bệnh (do virus, vi khuẩn, ký sinh trùng...) cho kết quả rất chính xác, di truyền học để sản xuất các marker phân tử, nghiên cứu sự phát sinh các đột biến..., đấu tranh chống tội phạm, nghiên cứu phát hiện mối quan hệ giữa người chết với người sống hoặc giữa các nhóm dân tộc... Mặt khác, việc phân tích trình tự và thành phần nucleotid của DNA có giá trị rất lớn trong việc định loại các loài sinh vật.
Nguồn tham khảo:
Trần Quốc Dung, VOER.
Nguồn tham khảo:
Trần Quốc Dung, VOER.






Không có nhận xét nào:
Đăng nhận xét